


![]() IPM Florida
IPM Florida
IPM Florida Personnel
Purpose of Website
IPM Florida Documents
IPM Projects
IPM Florida Partners
IPM Funding
IPM-Related Organizations
IPM Florida ListServ
![]() Extension/Education
Extension/Education
Extension Resources
IPM Education and Training
Photo Galleries
Presentations
Publications
Videos
Video Classics
![]() Agricultural IPM
Agricultural IPM
Forestry
Field Crops
Greenhouse Crops
Vegetable Crops
Fruit Crops
Ornamentals
Pastures, Turf and Sod
Livestock
Organic and Sustainable
Other Crops
![]() Community IPM
Community IPM
Biting, Stinging, Nuisance Pests
Household IPM
Landscape IPM
Home Gardening IPM
Public Health IPM
Institutional IPM
![]() Environmental IPM
Environmental IPM
Invasive Arthropods
Invasive Diseases
Invasive Plants
Invasive Species
![]() Applying IPM
Applying IPM
IPM Planning/Guides
Scouting
Pest Identification and Diagnosis
Pest Management Methods
IPM Adoption
Thrips IPM Program
Insect Rearing
Areawide IPM and SIT
![]() Additional Information
Additional Information
Southern States IPM Websites
EDIS
Crop Profiles
Pest Management Strategic Plans
Solutions for your Life
eXtension
Chinese privet, Ligustrum sinense, Prospects for Classical Biological Control in the Southeastern United States
 J. P. Cuda and M. C. Zeller
J. P. Cuda and M. C. Zeller
Jim Cuda is an assistant professor in the Entomology and Nematology
Department, University of Florida, P.O. Box 110620, Gainesville,
Florida 32611-0620
Mark Zeller, is a biologist in the Bureau of Invasive Plant Management,
Florida Department of Environmental Protection, 3917 Commonwealth
Boulevard, M. S. 710, Tallahassee, Florida 32399-3000.
Chinese or hedge privet, Ligustrum sinense (Oleaceae), is a semi deciduous shrub or small tree of Asiatic origin that is commercially available from the nursery industry as an ornamental or border plant (PlantFinder 1999). After its introduction into the United States in 1852 (Dirr 1983), this woody shrub eventually escaped cultivation, and by 1932 had become naturalized across the southeastern United States (Small 1933). It is now considered an invasive weed in many areas of the Southeast (Dirr 1983, Nelson 1996).
 In its introduced range, Chinese privet is a common component of
the understory vegetation of forests in north Florida (Fig. 1), Alabama,
Georgia (Godfrey 1988), Kentucky, North and South Carolina (Radford
et al. 1968), Mississippi (Goddard 1992), Oklahoma (Taylor et al.
1996), Tennessee (Radford et al. 1968, Faulkner et al. 1989) and
Texas (Correll and Johnston 1970). This woody shrub also is an invasive
weed in Australia (Burrows and Kohen 1986) and Argentina (Montaldo
1993). While primarily a woody invader of abandoned or disturbed
areas (Godfrey 1988), Chinese privet readily invades natural areas
adjacent to disturbed sites, and is especially abundant along stream
banks. The present distribution of naturalized populations of Chinese
privet in Florida is shown in Fig. 2 (Wunderlin et al. 1995).
In its introduced range, Chinese privet is a common component of
the understory vegetation of forests in north Florida (Fig. 1), Alabama,
Georgia (Godfrey 1988), Kentucky, North and South Carolina (Radford
et al. 1968), Mississippi (Goddard 1992), Oklahoma (Taylor et al.
1996), Tennessee (Radford et al. 1968, Faulkner et al. 1989) and
Texas (Correll and Johnston 1970). This woody shrub also is an invasive
weed in Australia (Burrows and Kohen 1986) and Argentina (Montaldo
1993). While primarily a woody invader of abandoned or disturbed
areas (Godfrey 1988), Chinese privet readily invades natural areas
adjacent to disturbed sites, and is especially abundant along stream
banks. The present distribution of naturalized populations of Chinese
privet in Florida is shown in Fig. 2 (Wunderlin et al. 1995).
Chinese privet can grow to a height of 10 m. The plant is characterized by its numerous leafy pubescent branchlets, and its small, simple, deciduous to semi deciduous leaves are, elliptic in shape (Fig. 3). The leaves are green (variegated in cultivation), opposite, have short pubescent petioles, and are attached at right angles to the stems. In bloom, the branchlets bear numerous, many flowered panicles. Flowers (Fig. 3) are white and malodorous; the corolla tube is shorter than the spreading lobes, and the stamens are exerted. The fruits (Fig. 4) are blue, fleshy drupes that are ingested by birds (McRae 1980, Buchanan 1989, Montaldo 1993). New infestations probably develop from bird dispersed seeds (Montaldo 1993).
The Florida Exotic Pest Plant Council lists Chinese privet as a Category I invasive species (FLEPPC 1996). The shrub disrupts native plant communities in riparian and upland habitats by competing for the available light and space. The federal and state listed endangered Miccosukee gooseberry, Ribes echinellum, is on the verge of being displaced in Florida by encroaching stands of Chinese privet (Langeland and Burks 1998). Moreover, nutrient cycling in natural areas can be disrupted by the presence of Chinese privet. Faulkner et al. (1989) observed that dense thickets of Chinese privet in Tennessee produce large quantities of litter, and act like umbrellas by preventing the infiltration of leaf litter from the native tree canopy. Agriculture and public health also can be impacted by the presence of Chinese privet. Dense stands of this invasive shrub can serve as alternate host plants for the citrus whitefly Dialeurodes citri (Hicks and Oliver 1987), and are conducive to infestations of the hard tick Ixodes scapularis, a suspected vector of Lyme disease in the southern United States (Goddard 1992).
Where it displaces native vegetation, Chinese privet may not have an adverse impact on wildlife populations because of its nutritional value. The plant serves as food source for the bobwhite quail, Colinus virginianus, and the white tailed deer, Odocoileus virginianus, when more desirable forage plants are scarce (McRae 1980, Stromeyer et al. 1998a,b).
From a management perspective, mechanical control methods such as cutting and prescribed burning are ineffective against Chinese privet (Faulkner et al. 1989). The plant resprouts rapidly from the cut stumps, and its affinity for low lying damp areas does not allow fire to carry well. The moist conditions in the bottom litter layer also reduce the effectiveness of the fire by not allowing temperatures to become hot enough to kill the root crowns. Although the herbicide glyphosate will severely damage or kill Chinese privet, chemical control is impractical and too expensive for large areas with dense growth (Faulkner et al. 1989). Since conventional methods are inappropriate for selectively controlling large infestations of Chinese privet in natural areas, biological control with natural enemies from the plant's native range should be considered.
Chinese privet would be a good candidate for classical biological control because the environmental risks associated with the importation and release of host specific natural enemies would be low. There are no native congeners in the genus Ligustrum occurring in the United States (Pemberton 1996). Therefore, the level of host specificity required for candidate natural enemies may be quite broad yet still not pose a risk to non target native or economically important plant species. The nursery industry would probably object to the introduction of insects that feed on the foliage or girdle the branches or stems of Chinese privet and would be fearful of attacks on the widely used Ligustrum japonicum (Japanese privet) and the also popular L. lucidum (wax privet). This conflict of interest could be resolved by selecting only those insects that attack the plant's flowers or seeds. According to Harley (1986), suppression of reproduction is important for controlling woody weeds (a) that reproduce by seed; (b) where existing plantings provide some benefit, e.g., ornamental value; and (c) when herbicide applications provide only temporary control and retreatment is uneconomical or environmentally disruptive. Chinese privet meets all of these criteria.
Biological control of Chinese privet already may be occurring in some areas, although not as a result of any purposeful introductions. The seed weevil Ochyromera ligustri was recently found attacking Chinese privet in Leon County, Florida (Fig. 2) (Cuda and Zeller 1998). This immigrant natural enemy of Ligustrum spp. may be capable of reducing the spread of Chinese privet into new areas and/or the densities of existing stands. However, field and laboratory studies will be required to determine to what extent this insect can control the growth and spread of Chinese privet populations in natural areas.
 Ochyromera ligustri was first discovered in 1959 on Japanese privet,
L. japonicum, in Wake County, North Carolina (Warner 1961, Wray 1961),
and is believed to have immigrated from the Orient in nursery stock
imported into the United States. Since then, the weevil has been
found in Florida, Georgia, North and South Carolina, and Virginia,
as well as North and South Dakota (O'Brien and Wibmer 1982, Johnson
and Lyon 1988). Considering that Leon County is the only locality
in Florida where Japanese privet (the principal host plant for O.
ligustri) is documented as naturalized (Wunderlin et al. 1995), it
is not surprising that the weevil has adapted to Chinese privet in
this region of north Florida where the two species of Ligustrum have
overlapping distributions. The weevil also has been collected from
wax leaf ligustrum, L. lucidum, common privet, L. amurense, lilac,
Syringa spp., and grape, Vitis spp., in North Carolina (Warner 1961,
Wray 1961). However, it is not clear from the literature whether
lilac or grape are legitimate host plants in that they are capable
of supporting complete development of the weevil.
Ochyromera ligustri was first discovered in 1959 on Japanese privet,
L. japonicum, in Wake County, North Carolina (Warner 1961, Wray 1961),
and is believed to have immigrated from the Orient in nursery stock
imported into the United States. Since then, the weevil has been
found in Florida, Georgia, North and South Carolina, and Virginia,
as well as North and South Dakota (O'Brien and Wibmer 1982, Johnson
and Lyon 1988). Considering that Leon County is the only locality
in Florida where Japanese privet (the principal host plant for O.
ligustri) is documented as naturalized (Wunderlin et al. 1995), it
is not surprising that the weevil has adapted to Chinese privet in
this region of north Florida where the two species of Ligustrum have
overlapping distributions. The weevil also has been collected from
wax leaf ligustrum, L. lucidum, common privet, L. amurense, lilac,
Syringa spp., and grape, Vitis spp., in North Carolina (Warner 1961,
Wray 1961). However, it is not clear from the literature whether
lilac or grape are legitimate host plants in that they are capable
of supporting complete development of the weevil.
 The biology of O. ligustri on Japanese privet was reviewed by Johnson
and Lyon (1988). The weevil is 3 5 mm long and is shiny brown with
golden yellow hair like scales (Fig. 5). The adults feed on the foliage
by making small perforations in the interior of the leaf blade, and
drop to the ground when disturbed. In North Carolina, adults are
active from late June to early July. Females deposit their eggs in
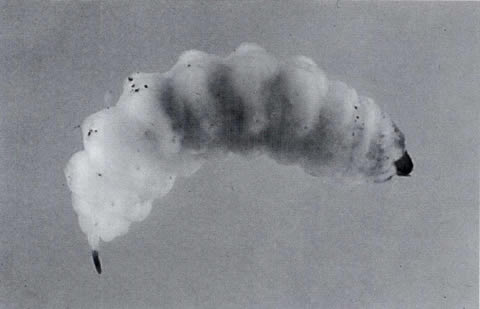
the seed or in the mature fleshy fruits. The larvae (Fig. 6) feed
inside the fruits and destroy the seeds as they complete their development
to the adult stage. New adults emerge from the seeds (one weevil
per seed) in mid May, and there is only one generation per year.
The biology of O. ligustri on Japanese privet was reviewed by Johnson
and Lyon (1988). The weevil is 3 5 mm long and is shiny brown with
golden yellow hair like scales (Fig. 5). The adults feed on the foliage
by making small perforations in the interior of the leaf blade, and
drop to the ground when disturbed. In North Carolina, adults are
active from late June to early July. Females deposit their eggs in
the seed or in the mature fleshy fruits. The larvae (Fig. 6) feed
inside the fruits and destroy the seeds as they complete their development
to the adult stage. New adults emerge from the seeds (one weevil
per seed) in mid May, and there is only one generation per year.
Acknowledgements
We thank M. C. Thomas for identifying the insect, J. F. Butler and K. C. Burks for the photographs of the insect and plant, respectively, and J.H. Frank and N.C. Coile for reviewing the manuscript. Florida Agricultural Experiment Station Journal Series No. N 01574.
References
Buchanan, R. A. 1989. Pied currawongs (Strepera graculina): Their diet and role in weed dispersal in suburban Sydney, New South Wales (Australia). Proceedings of the Linnaen Society of New South Wales 111: 241 256.
Burrows, F. J., and J. Kohen. 1986. Inhibition of germination in privet. Plant Protection Qtrly 1:107 108.
Correll, D. S., and M. C. Johnston. 1970. Manual of the vascular plants of Texas.Texas Research Foundation, Renner, Texas. 1881 pp.
Cuda, J. P., and M. C. Zeller. 1998. First record of Ochyromera ligustri (Coleoptera: Curculionidae) from Chinese privet in Florida. Florida Entomologist 81: 582 584.
Dirr, M.A. 1983. Manual of woody landscape plants, 3rd ed. Stipes Publishing Co., Champaign, IL. 536 pp.
Faulkner, J. L., E. E. C. Clebsch, and W. L. Sanders. 1989. Use of prescribed burning for managing natural and historic resources in Chickamauga and Chattanooga National Military Park, USA. Environmental Management 13: 603 612.
Florida EPPC. 1996. Florida Exotic Pest Plant Council occurrence database. URL: http:// www.fleppc.org/.
Goddard, J. 1992. Ecological studies of adult Ixodes scapularis in central Mississippi: questing activity in relation to time of year, vegetation type, and meteorologic conditions. Journal of Medical Entomology 29: 501 506.
Godfrey, R. K. 1988. Trees, shrubs, and woody vines of northern Florida and adjacent Georgia and Alabama. University of Georgia Press, Athens. 734 pp.
Harley K. L. S. 1986. Suppression of reproduction of woody weeds using insects which destroy flowers or seeds, pp. 749 756. In Delfosse, E.S. (ed.), Proceedings of the VI International Symposium on Biological Control of Weeds, 19 25 August 1984, Vancouver, Canada. Agriculture Canada.
Hicks, C. B., and A. D. Oliver. 1987. Citrus whitefly preference among four ornamental host plants. Hortscience 22: 59 60.
Johnson, W. T., and H. H. Lyon. 1988. Insects that feed on trees and shrubs, 2nd ed. Cornell University Press, Ithaca. 556 pp.
Langeland, K. A., and K. C. Burks. 1998. Identification and biology of non-native plants in Florida's natural areas. University of Florida, Gainesville. 165 pp.
McCrae, W. A. 1980. Unusual bobwhite foods on abandoned Piedmont farmlands, Georgia, USA. Georgia Journal of Science 38: 49 54.
Montaldo, N. H. 1993. Avian dispersal and reproductive success of two species of Ligustrum (Oleaceae) in a forest relict in Argentina. Revista Chilena de Historia Natural 66: 75 85.
Nelson, G. 1996. The shrubs and woody vines of Florida. Pineapple Press, Sarasota. 391 pp.
O'Brien, C. W., and G. J. Wibmer. 1982. Annotated checklist of the weevils (Curculionidae sensu lato) of North America, Central America, and West Indies (Coleoptera: Curculionoidea). Memoirs of the American Entomological Institute 34: 116.
Pemberton, R. W. 1996. The potential of biological control for the suppression of invasive weeds of southern environments. Castanea 61:313 319.
PlantFinder. 1999. PlantFinder wholesale guide to foliage and ornamental plants, October, p. 165.
Radford, A. E., H. E. Ahles, and C. R. Bell. 1968. Manual of the vascular flora of the Carolinas. University of North Carolina Press, Chapel Hill. 1183 pp.
Small, J. K. 1933. Manual of the southeastern flora, part one and two. University of North Carolina Press, Chapel Hill. 1554 pp. (facsimile reprint 1972, Hafner Publishing, New York.).
Stromayer, K. A. K, R. J. Warren, and T. B. Harrington. 1998b. Managing Chinese privet for white tailed deer. Southern Journal of Applied Forestry 22: 227 230.
Stromayer, K. A. K, R. J. Warren, A. S. Johnson, P. E. Hale, C. L. Rogers, and C. L. Tucker. 1998a. Chinese privet and the feeding ecology of white tailed deer: The role of an exotic plant. Journal of Wildlife Management 62: 13211329.
Taylor, E. S., L. K. Magrath, P. Folley, P Buck, and S. Carpenter. 1996. Oklahoma vascular plants: Additions and distributional comments. Proceedings of the Oklahoma Academy of Sciences 76: 31 34.
Warner, R. E. 1961. The genus Ochyromera new to the western hemisphere, with a new species and additions to the Junk Schenkling Coleopterorum Catalogus (Curculionidae: Prionomerinae, Endaeini). Coleopterists' Bulletin 15: 121 124.
Wray, D. L. 1961. Biology and life history of the ligustrum weevil (Curculionidae). Coleopterists' Bulletin 15: 119 120.
Wunderlin, R. P., B. F. Hansen, and E. L. Bridges. 1995 (updated Dec 1998). Atlas of Florida vascular plants. University of South Florida, Institute for Systematic Botany.